En Ikke-Tilfeldig Evolusjons-Hypotese (EITEH)
(oversatt fra 'The Evolution Revolution' Kap. 2 -av Dr. Lee M. Spetner, Jerusalem, Israel, Juni 2014.)
Epigenetikk er blitt kalt et av de nye yndlingsordene i biologien (Jablonka, 2009). Termen ble først introdusert i 1942 for å beskrive arvelige endringer i genetisk funksjonsevne, i løpet av embryo-perioden, men termen innebærer mer enn det. Et epigenetisk trekk er ofte definert som et arvelig trekk, som resulterer i en genetisk endring uten å innebære endring i DNA-sekvensen (Jablonska, 2009). Termen brukes spesielt om genetiske endringer i løpet av embryo-perioden, som skyldes input fra nabo-celler i løpet av utvikling av embryoet. Men denne bruksmåten ekskluderer det J. Shapiro har kalt naturlig genetisk engineering (1992, 1997, 2005). Epigenetiske endringer innbefatter alle typer av genetiske endringer som er stimulert av omgivelsene, inkludert de som kommer fra utsiden av organismen.
Bilde 1.  En Ikke-Tilfeldig Evolusjons-Hypotese
En Ikke-Tilfeldig Evolusjons-Hypotese
I boka 'Not by Chance' introduserte forfatteren at mye av evolusjonen vi faktisk observerer, er resultat av organismens innebygde kapasitet til å gi tilpassede svar til input fra omgivelsene. Forfatteren kalte det En Ikke-Tilfeldig Evolusjons-Hypotese (EITEH). Denne type endringer støtter seg til begivenheter som er epigenetiske på bredt grunnlag. Det skiller seg merkbart fra Neo-Darwinsk evolusjon, som postulerer at en populasjon endrer seg når en sjelden tilfeldig mutasjon brer seg over populasjonen, gjennom naturlig seleksjon. Neo-Darwinsk evolusjon sies å være drevet av tilfeldige genetiske endringer, mens den type evolusjon forfatteren forslår er drevet av ikke-tilfeldige epigenetiske endringer, som trigges av input fra omgivelsene.
Forfatteren har foreslått at en endring i omgivelsene kan forårsake genomet til et individ å endres, for å sette i verk et tilpasset svar til endringen, og dette endrede genomet kan arves. Det er kjent generelt at endringer i omgivelsene kan stimulere epigenetiske endringer, men det er ikke så kjent at en betydelig andel av disse er tilpasset til omgivelsene som utførte stimuleringen. Dyr og planter har en innebygde evne til å reagere tilpasset til omgivelsenes stimuli. Denne evnen setter plantene og dyrene i stand til raskt å tilpasse seg til endrede omgivelser.
Ordet tilfeldig blir ofte brukt når det refereres til mutasjoner som er kilde til nyheten i neo-darwinsk evolusjonsteori. Ordet 'tilfeldig' innebærer at disse mutasjonene inntreffer uavhengig av den effekt de har på arten (fenotypen). Det er i denne betydning at mutasjoner sies å være tilfeldige i evolusjonsteorien (1). Det er denne statistiske uavhengigheten som har tjent til å kjennetegne den neo-darwinske evolusjonsteorien fra hypoteser om arv av oppnådde egenskaper, hvorav den mest kjente var Lamarckismen. Selv om ikke Darwin selv benyttet ordet tilfeldig, så har han blitt forstått å ha ment det, når han ikke prøvde å inkludere Lamarcks ideer.
Dr. Spetner foreslo 'En Ikke-Tilfeldig Evolusjons-Hypotese' (EITEH), fordi den kunne redegjøre for raske evolusjonære endringer som har funnet sted. Evnen til å gi tilsvar til endringer i omgivelsene, krever en organisme som er i stand til å oppfatte en endring i omgivelsene og har en mekanisme hvorved denne sansede endringen fører til aktivitet av et latent gen, eller andre genetiske ressurser. Disse vil i sin tur lede til endring i fenotypen, som gir organismen en fordel i de nye omgivelsene. B. Sanner ved Emory Universitet har foreslått noe likt dette for bakterier, at genomisk rearrangement kan trigges av endringer i omgivelsene via et kontrollsystem i bakterien som kan gi arvelige endringer (Wanner, 1985). Chr. Cullis og kolleger ved CaseWestern Reserve Universitet i Cleveland har rapportert at miljøendringer framkaller genetisk rearrangement i linplanter. Det er de samme genetiske endringene som inntreffer når de samme miløendringer inntreffer, noe som indikerer at genetiske endringer stimuleres av omgivelsene ikke ved tilfeldigheter. (Cullis 2005, Chen et al. 2009)
Siden de samme endringene inntraff i hvert individ under samme forhold, mente Cullis at det var fornuftig å foreslå at endringene er tilpassede.
A. Prevosti og kolleger ved Barcelona Universitet har funnet at layout til DnA i bananfluen (Drosophila subobscura) varierer med lengdegrad. Samme variasjon er funnet både i Nord-Amerika og Europa (Prevost et al. 1988). Hvordan kan dette forstås? Variasjonen i DNA kan bli forstått som det mulige resultat av epigenetisk respons mot omgivelsene. Likheten i variasjonen i DNA på to kontinent indikerer at i hvert av tilfellene er DNA-layout relatert til miljømessige forhold, som er karakteristiske, og er gjensvar på breddegraden de inntreffer under. Tilsynelatende har input fra lokalomgivelsene vært ansvarlige for genetisk rearrangering i bananfluer (Lee 2002).
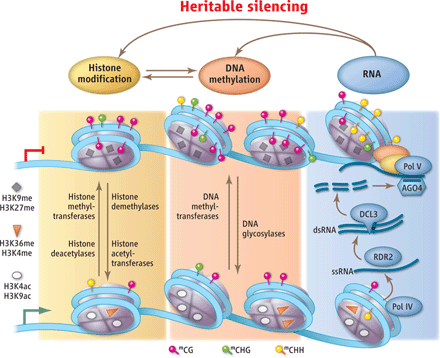 Forfatteren foreslo i sin tidligere bok ('Not by Chance') at en celle, og t.o.m. en organisme kunne ha en innebygd evne til å å utøve langtids-kontroll, som kunne vare over generasjoner, ved å tilpasse seg langtids-endringer. Fra forskningsrapporter som har kommet siden da, er dette forslaget blitt rettferdiggjort. En miljømessig endring kan bli en langtids-utfordring og organismen kan svare gjennom arvelige endringer, som tjener til å tilpasse den og dens avkom. Cellen kan ha andre trick for å fullføre samme hensikt. Denne evnen har en likhet med dets evne til å utøve korttidst-kontroll.
Forfatteren foreslo i sin tidligere bok ('Not by Chance') at en celle, og t.o.m. en organisme kunne ha en innebygd evne til å å utøve langtids-kontroll, som kunne vare over generasjoner, ved å tilpasse seg langtids-endringer. Fra forskningsrapporter som har kommet siden da, er dette forslaget blitt rettferdiggjort. En miljømessig endring kan bli en langtids-utfordring og organismen kan svare gjennom arvelige endringer, som tjener til å tilpasse den og dens avkom. Cellen kan ha andre trick for å fullføre samme hensikt. Denne evnen har en likhet med dets evne til å utøve korttidst-kontroll.
J. Shapiro har foreslått at celler har evne til å utføre sin egen genetiske engineering (Shapiro 1997, 1999, 2011). Denne evnen er innebygd i cellen, og setter organismen i stand til å endre sitt genom for å tilpasse seg endrede omgivelser. Organismer har således evnen til raskt å tilpasse seg nye omgivelser. Vi har ingen teori for hvordan denne evnen ble integrert i organismen. De evolusjonære begivenhetene som har blitt direkte observert, kan ikke gjøre rede for det. Heller ikke kan de gjøre rede for felles avstamning. Det er ingen kjent prosess gjennom hvilken den nødvendige informasjon kunne ha blitt bygd opp. Det kan dermed ikke bevises at utvikling av livet, nødvendigvis skjer gjennom en overnaturlig begivenhet, men det viser tomheten ved nåværende teorier om opprinnelsen og utviklingen av livet, gjennom felles avstamning.
Det har skjedd mye læring om hvordan organismer kan endre sitt genom de siste tiårene. Flyttbare genetiske elementer ble oppdaget under 2.vkrig av B. McClintock (McClintlock, 1941, 1950, 1955, 1983), men ble i starten avvist som uekte fenomen. Selv om det ble ansett som dødfødt, fortsatte McClintock sin forskning og omsider ble viktigheten av arbeidet anerkjent ved at hun fikk Nobelprisen i Medisin i 1983. De flyttbare genetiske elementene hun oppdaget, har senere vist seg å være medlemmer av en klasse genetiske rearrangement, som ikke inntreffer spontant tilfeldig, men er under streng cellulær kontroll. Seksjoner av DNA kan flyttes fra ett sted i genomet til ett annet, eller kan fjernes helt. Disse kontrollerte genetiske endringene kan vise skjulte, latente gener som var i genomet, men tidligere var utilgjengelige for organismen. Vi skal se mer senere på disse kryptiske genene, som Hall (1999) kaller dem.
Bilde 2. Arvelige endringer -uten endring i DNA-sekvens
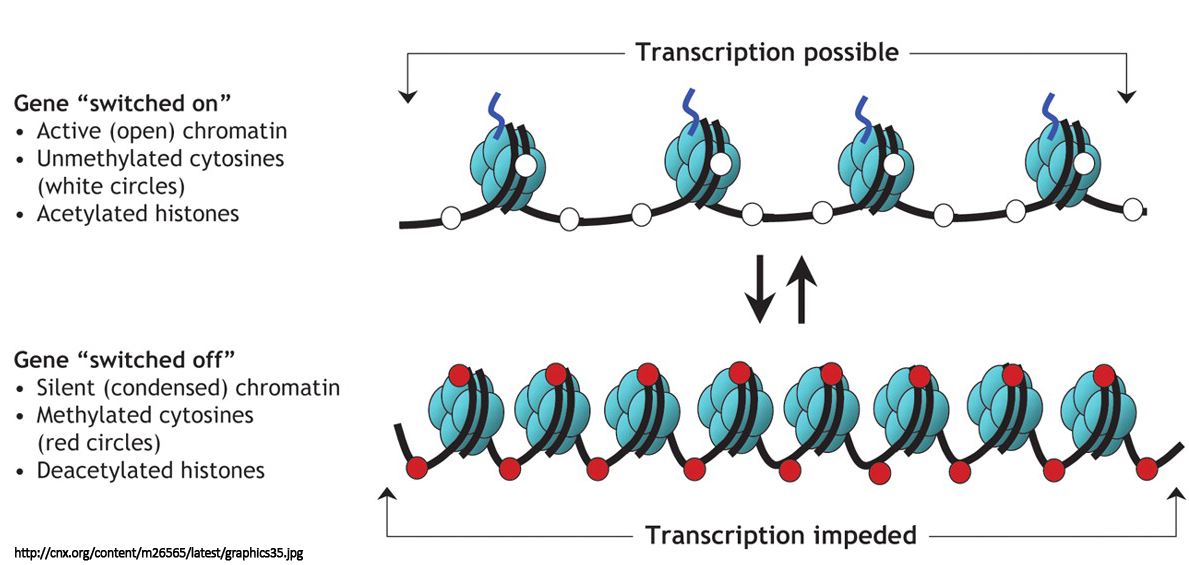 Endringer i omgivelsene vites å få fram ulike typer stress-reaksjoner i en organisme. McClintock viste i sitt tidlige arbeid med planter, at noen typer stress kan trigge genetiske rearrangeringer (McClintock, 1984). Organismer synes å ha evnen til å lindre stresset, ved å endre både sin feno- og sin geno-type. Stress er generelt definert som en miljømessig betingelse, som truer med å velte balansen og stabiliteten i organismen. Stress i en organisme inkluderer mange slags stimuli. I mikroorganismer kan stress føre til en overflod eller berøvelse av nødvendige molekyler, slik som sukker- eller salt-typer. Det kunne også bli ekstremt høye eller lave temperaturer. I planter og dyr, så er stress vanligvis en mer kompleks form for angrep fra omgivelsene. Stress kan frambringe genetiske rearrangement, som i sin tur kan aktivere skjulte (kryptiske) gener. Det er kjent mange eks. omkring dette (Shapiro 1992 & 2009, Hall 1999). Slack et al. har (2006) rapportert at stress kan frambringe en tilpasset respons i E.Coli ved en selektiv forsterkerkrets av gener. Hersh et al (2004) har rapportert at stress kan forårsake tilpassede genendringer i Ecoli. Vi skal se på både encellede og høyere organismer, samt arvelig effekt hos disse i det videre.
Endringer i omgivelsene vites å få fram ulike typer stress-reaksjoner i en organisme. McClintock viste i sitt tidlige arbeid med planter, at noen typer stress kan trigge genetiske rearrangeringer (McClintock, 1984). Organismer synes å ha evnen til å lindre stresset, ved å endre både sin feno- og sin geno-type. Stress er generelt definert som en miljømessig betingelse, som truer med å velte balansen og stabiliteten i organismen. Stress i en organisme inkluderer mange slags stimuli. I mikroorganismer kan stress føre til en overflod eller berøvelse av nødvendige molekyler, slik som sukker- eller salt-typer. Det kunne også bli ekstremt høye eller lave temperaturer. I planter og dyr, så er stress vanligvis en mer kompleks form for angrep fra omgivelsene. Stress kan frambringe genetiske rearrangement, som i sin tur kan aktivere skjulte (kryptiske) gener. Det er kjent mange eks. omkring dette (Shapiro 1992 & 2009, Hall 1999). Slack et al. har (2006) rapportert at stress kan frambringe en tilpasset respons i E.Coli ved en selektiv forsterkerkrets av gener. Hersh et al (2004) har rapportert at stress kan forårsake tilpassede genendringer i Ecoli. Vi skal se på både encellede og høyere organismer, samt arvelig effekt hos disse i det videre.
Bilde 3. Gener kan slås av og på
Den ikke-tilfeldige evolusjonære hypotesen som foreslås her (EITEH), er meget ulik den om tilfeldige mutasjoner i neo-darwinismen. Forfatteren foreslår en evolusjonær prosess i hvilken individer utvikler seg, motsatt neo-darwinismens prosess der populasjoner endrer seg. Stress kan iverksette genetiske endringer i en organisme, som kan aktivere et sett av latente gener i både somatiske celler og kjønnscellene. Dersom omgivelsene endres signifikant, vil stressreaksjoner på dette medføre genetiske rearrangement. Det tilpassede svaret er en egenskap innebygd i organismen, for å matche kilder til stress i omgivelsene. De genetiske endringene ville kunne forekomme i hele populasjonen eller bare i en stor andel av den. Om tilpassede endringer skjer i kjønnscellene, vil snart hele populasjonen bestå av individer med endret genetikk, som følge av naturlig seleksjon. (2) Naturlig seleksjon kan være effektiv når endringer forekommer i mange individer, men kan ikke grunngi felles avstamning.
Genetiske rearrangement blir verksatt gjennom repetive seksjoner i DNA (Shaphiro og von Sternberg 2005, Shaphiro 2011). De viktigste DNA-elementer for rearrangering er transposoner, som består av integrerte system av proteiner og aminosyrer. Transposoner, som en før kalte 'hoppende gener' kan endre posisjon i genomet. De kan, i tillegg til å hoppe omkring i genomet selv, kontrollere bevegelse til andre segmenter i genomet. De kan replikere seg selv (copy-paste) og plassere kopien annensteds i genomet. Eller de kan bare flytte (cut and paste) uten å replikere seg selv (Shaphiro, 1999). Disse elementer er de mest tallrike i menneskelig genom. Mer enn 42% av menneskelig genom består av dem (Smit 1999). OM en sammenligner med hyppigheten til protein-kodende DNA, som kun utgjør ca. 2 % av menneskelig genom. Videre er denne type av repeterende DNA spesifikk for hver kategori av organismer, og skiller seg enda mer enn protein-kodende deler av DNA. I pattedyr er spesielt SINE(Short-Interperced-Nucleotide-Elements)-DNA, som er ansvarlig for deler av DNA-arrangement, kjent for å være unik for hver orden av pattedyr (Shaphiro 2002).
Ikke-tilfeldige genetiske endringer, trigget av omgivelses-Input, kan gjøre rede for eksempler av evolusjon som faktisk har blitt observert (i motsetning til 'konkludert'). Det samme gjelder ikke for tilfeldige mutasjoner, som følge DNA kopierings-feil. Genetiske rearrangeringer kan produsere genetiske forskjeller mellom nært beslektede organismer. En celle har evnen til å flytte rundt deler av genomet, ved å undertrykke gener som ikke trengs og åpenbare de som tidligere var skjult, og har mulighet til å tilpasse organismen til nåværende omgivelser. Videre er de genetiske rearrangement, som vil få fram tilpassede gener, kjent for å kunne trigges av omgivelsene.
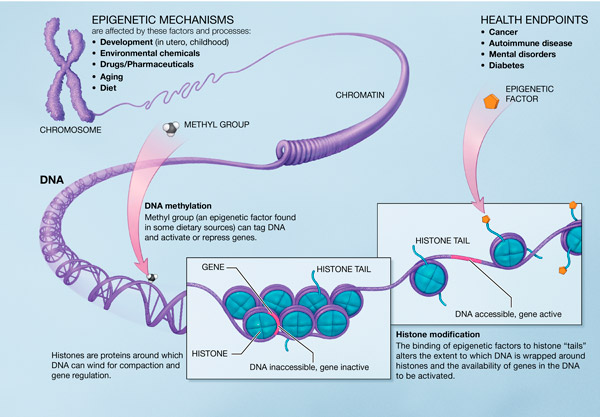
Bilde 4. Epigenetetiske mekanismer
En organisme har dermed en innebygd evne til å tilpasse seg til nye omgivelser ved å endre sitt DNA. Disse tilpasningene inntreffer bare når de behøves, fordi de er trigget av input fra de nye omgivelsene. Siden det er omgivelsene som frambringer dem, er de ikke sjeldne i bestanden. De vil framstå i en stor del av populasjonen, og lede til raske evolusjonære endringer. Muligens endog innen én generasjon. Om det samme skulle skje via tilfeldige punktmutasjoner, måtte det skje over lange tidsperioder, om de i det hele kunne gjennomføres. Det er ikke en gang kjent om det eksisterer noen slik farbar vei, der hver lille endring fra forrige generasjon ville føre til en bedre tilpasning for neste generasjon. Neo-darwinister formoder alltid at slike finnes, men har ikke bevist at det gjør det.
Epigenetiske endringer forårsaket av miljøet, er ikke-tilfeldige og er forskjellig fra tilfeldige punkt-mutasjoner. Epigenetiske endringer kan komme til syne i mange individer, som erfarer endrede omgivelser. De vil meget sannsynlig øke i antall og den nye genetiske endringen, ville være et kjennetegn ved populasjonen. Enkeltvise punkt-mutasjoner vil meget sannsynlig gå tapt fra populasjonen, selv om de i prinsippet kan ha positiv selektiv verdi (Kap.1).
Kommentarer:
1. Evolusjonister kan også si at ordet tilfeldig betyr uniformt fordelt. Når de sier at mutasjonsrater er tilfeldige, mener de ofte at mutasjonsrater er like store, og om de ikke er like store, sier de at de ikke er tilfeldige. Men den bruken er ikke i samsvar med ordet tilfeldig i matematikk.
2. Forfatteren er positiv i forhold til naturlig seleksjon, men avviser felles avstamning, som ikke kan gjøres rede for ved naturlig seleksjon. Naturlig seleksjon kan være meget effektiv, når tilpassede genetiske endringer opptrer i mange individer -i stedet for bare ett.
Stoffutvalg og bilder ved Asbjørn E. Lund